
認知・情動脳科学専攻 Major of Cognitive and Emotional Neuroscience
Evolution of habenular asymmetry/lateralization in vertebrates
 解剖学・神経科学 Anatomy and Neuroscience
解剖学・神経科学 Anatomy and Neuroscience一條 裕之 Hiroyuki Ichijo
- TEL : 076-434-7205
- URL : http://www.med.u-toyama.ac.jp/anat/
- Keywords : habenula, asymmetry, lateralization, natural selection, mouse, fish
研究の背景と目的 Background and Purpose of Study
神経回路の非対称性は脊椎動物で広く報告されており,それは2種類に分類することができます.一つは外界の空間的な非対称性と関連しており,非対称な行動を伴うものです.他方,外界の空間的な非対称性との関連が乏しく,非対称な行動を伴わないものがあります.その代表的な例が手綱核です.手綱核は間脳の背側に在り,終脳からの多様な入力を受け取り,中脳のモノアミン系神経回路に出力し,中枢神経系の全体を調節することが知られている神経核です(図1).脊椎動物において広く保存されているので,生存に必須な神経回路を構成していると考えられます.それに加えて,左右の手綱核の大きさが異なっており,非対称であることは20世紀のはじめから多く報告されてきました.
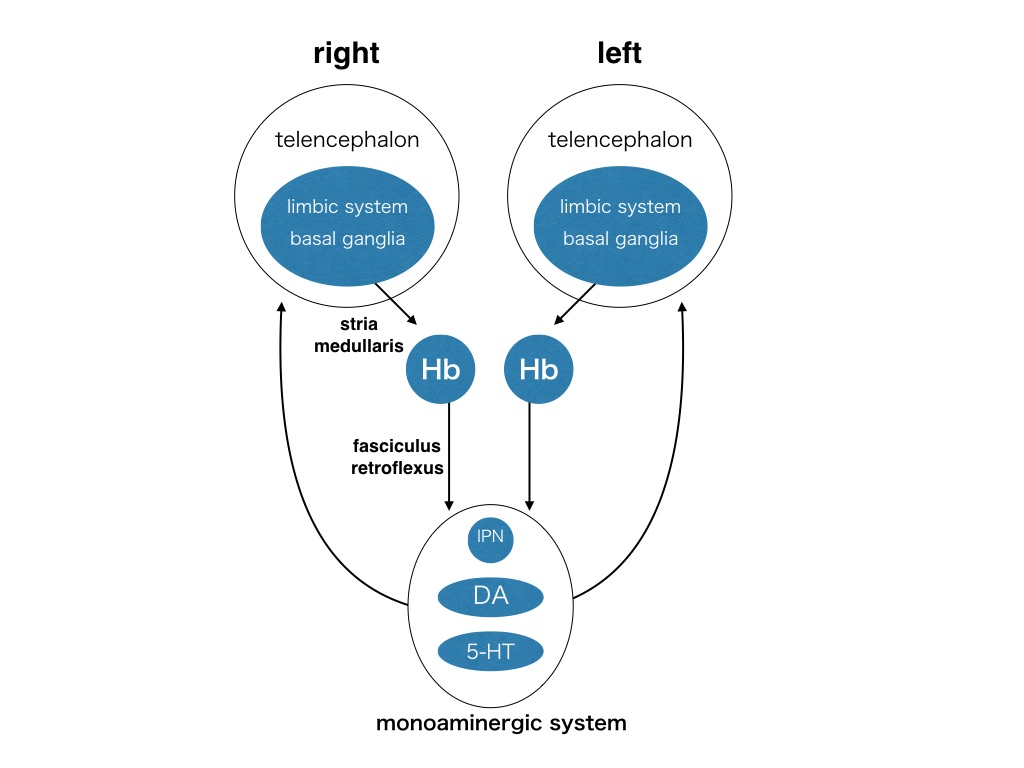
図1 手綱核の神経回路の模式図. 手綱核は広く情報をまとめて片側にまとめ,両側に拡げる情報のハブの位置にある.文献3)から引用
手綱核は覚醒(waking)と睡眠(sleeping )や概日リズム,報酬(reward)と罰(punishment),脅威(threat)に対する逃走行動(escaping)とすくみ行動(freezing),社会的な葛藤(social conflict,例えば喧嘩)における勝ち(winning )と負け(loosing),ストレス反応などに関わることが報告されています.手綱核は多様な現象との関わりが報告されている反面,それは雑然としており,その機能の本質をみえにくくしています.私達は手綱核の基本的な機能とその左右性がどの様に進化したかを探索しています.
Many vertebrates have asymmetrical circuits in the nervous system. There are two types of circuit asymmetry. One is related to environmental asymmetry in outer world and lateralized behaviors, but the other is with no relation to environmental asymmetry and lateralized behaviors. Typical example without lateralized behaviors is the habenula. The habenula is situated in dorsal diencephalon, receives various inputs from the telencephalon, sends outputs to mesencephalic monoaminergic modulation systems, and regulates various aspects of behaviors. It is reported variety of habenular function: circadian rhythm, waking and sleeping, reward and punishment, escaping and freezing under threats, winning and loosing under social conflict, stress responses, and so on. Because these are seemingly confused, one loses real nature of habenula. The habenula is conserved in vertebrates, indicating essential function in survival; moreover, it is well known that the habenula shows structural asymmetry in many vertebrate. Until recently, there had been no hypothesis regarding the evolution of habenular asymmetry. We proposed a hypothesis that binary opposition in functional incompatibility applies selection pressure on the habenular circuit and leads to asymmetry. We are investigating basic roles of the habenula and evolution of its asymmetry/lateralization.
本研究の領域横断性
研究においては標準的な実験を行うために特定の動物を利用することが普通です.たとえばマウス,ニワトリ,アフリカツメガエル,ゼブラフィッシュ,線虫,ショウジョウバエなどはモデル動物として神経科学の研究に大きな貢献をしてきましたし,これからもそうあり続けるでしょう.私達もその恩恵を受けて研究をしています.それとは別に,これまでに研究に利用されることが少なかった様々な動物は非モデル動物と呼ばれています.非モデル動物を利用して研究を行うことにも大きな意義があります.非モデル動物の中には特徴的な行動を示すものがあります.その背後にある機序を明らかにすることは特別な現象を理解することにとどまらず,神経回路機序の共通原理を明らかにする特異的な道筋になる事が期待されます. 私達はモデル動物と非モデル動物を分け隔てなく利用して,手綱核の左右性の進化を研究します.形態学的方法,生理学的方法,動物行動学的方法,分子遺伝学的方法を利用します.
研究内容
私達は報告されている多様な行動のそれぞれに手綱核が特異的に関わるのではなく,行動に含まれる共通要素に関わると考え,それぞれが機能的に両立しない二項対立行動であることを見いだしました.二項対立(binary opposition)はヒューリスティックに世界を定式化して情報を圧縮する機能を担うもので,広範にヒトの集団(部族)で認められる生得的な性質であると考えられています1).機能的に両立しない性質(functional incompatibility)が自然選択圧として働いたときに,並列した神経回路が進化することが指摘されており2),手綱核においても並列した回路が脳の右と左に割り当てられる結果,非対称な手綱核の神経回路が進化の過程で選択され,回路の全長が短くシンプルになり,左右に重複した同じ神経回路をもつことが避けられた結果,情報伝達の効率が上がり,適応度が高まったと考えられます(図2)3).このような理論的な枠組みの作成を出発点として,私達は手綱核の左右性の進化を通じて明らかにすることを目的としています.「機能的に両立しない二者択一的な行動選択仮説」を検証するために,魚類の行動の左右性と,手綱核の非対称性の相関を検証しています.
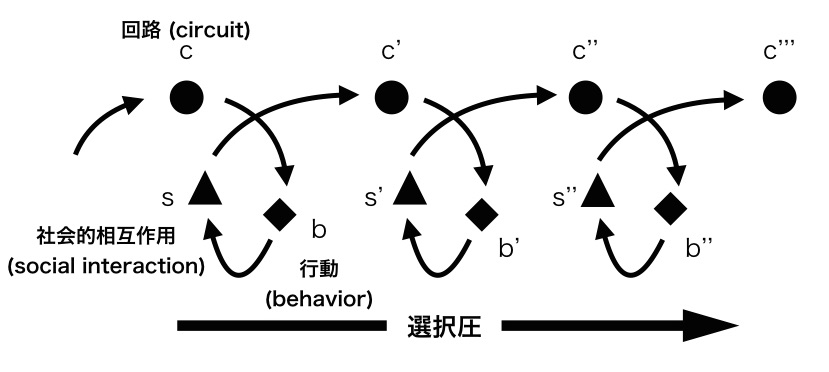
図2 神経回路と行動・社会性のらせん状の進化
マウスの手綱核の構造は対称ですが,非対称に機能することを私達は示しました(図3)4).マウスは複雑な環境の下で柔軟に情報を処理する機能的に非対称な回路を有していると考えられます.種間の進化戦略の違いを理解するために,マウスの手綱核の発達に伴った感受性の変化と,手綱核の左右性を導く上流の神経回路の探索を行っています.遺伝子・回路・個体の行動・集団の社会性の異なった水準を段階的に検証して,手綱核の非対称性の進化を理解することを試みています.
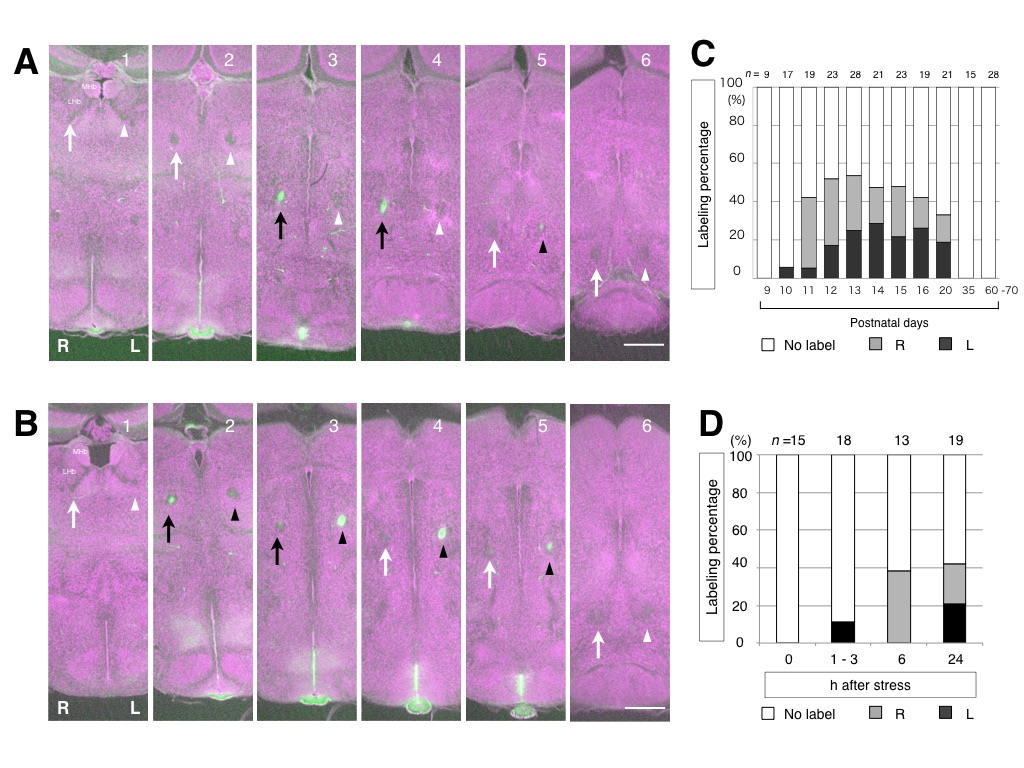
図3 マウスの手綱核の活動性の履歴の標識左右が非対称に活動することが示される.その活動性はストレス付加によって引き起こされ,幼若期に大きく,成熟すると穏やかになる.文献3)から引用
参考文献
- Lévi-Strauss C, Structuralism and Ecology. 1972
- Sherry DF, Schacter DL, The evolution of multiple memory systems. Psychol Rev, 94: 439-454. 1987.
- Ichijo H, Nakamura T, Kawaguchi M, Takeuchi Y. An evolutionary hypothesis of binary opposition in functional incompatibility about habenular asymmetry in vertebrates. Front Neurosci 10: 595. 2017.
- Ichijo H, Hamada M, Takahashi S, Kobayashi M, Nagai T, Toyama T, Kawaguchi M. Lateralization, maturation, and anteroposterior topography in the lateral habenula revealed by ZIF268/EGR1 immunoreactivity and labeling history of neuronal activity. Neurosci Res 95: 27-37. 2015.